



Effect of Different Levels of Social Isolation on the Acoustical Characteristics of Sheep Vocalization
Engeldal SEC1, Subandriyo2, Handiwirawan E3, Noor RR4
1Animal Science and Production Technology, Graduate School, Bogor Agricultural University
E-mail: angedevalle@yahoo.com
2Indonesian Institute for Animal Production
3Indonesian Center for Animal Research and Development
4Department of Animal Science and Production Technology, Bogor Agricultural University
(Diterima 3 Juni 2013 ; disetujui 20 Agustus 2013)
ABSTRAK
Engeldal SEC, Subandriyo, Handiwirawan E, Noor RR. 2012. Pengaruh tingkat isolasi sosial yang berbeda pada karakteristik akustik dari suara domba. JITV 18(3): 208-219. DOI: 10.14334/jitv.v18i3.323.
Pada hewan ternak, analisis vokal diterima sebagai metode non-invasif untuk menilai kesejahteraan hewan dibandingkan dengan pengukuran fisiologis. Penelitian ini bertujuan untuk meneliti pengaruh dari beberapa tingkat isolasi sosial pada karakteristik suara dari tiga bangsa domba yang berbeda. Domba diberi perlakuan yang terdiri dari tiga tingkat isolasi sosial yang berbeda dan selama perlakuan kemudian dicatat tingkah laku lokomotif dan suara domba perlakuan direkam. Data tingkah laku lokomotif dianalisa dengan metode deskriptif sedangkan software akustik digunakan untuk mengukur sebanyak tiga puluh enam parameter dari suara domba yang direkam. Analisa deskriptif menunjukkan bahwa domba dengan perlakuan isolasi parsial memperlihatkan tingkah laku lokomotif dan bersuara lebih banyak dibandingkan dengan isolasi lengkap. Analisa ANOVA menunjukkan bahwa tingkat isolasi dan bangsa domba mempunyai pengaruh nyata pada karakteristik struktur dan panjang suara. Hasil dari penelitian ini juga menunjukkan bahwa untuk karakteristik akustik amplitudo, kekuatan dan waktu mempunyai pengaruh kepada respon vokal selama isolasi. Karakteristik vokal frekuensi juga menunjukkan pengaruh yang nyata (P < 0,05) berdasarkan bangsa domba. Analisa spektrogram menunjukkan bahwa distribusi energi dalam suara domba adalah faktor yang paling jelas untuk membedakan di antara bangsa domba dan tingkat isolasi sosial. Hasil penelitian ini menunjukkan bahwa analisa akustik bisa bermanfaat untuk mengungkapkan keadaan afektif dari domba pejantan pada waktu isolasi sosial.
Kata Kunci: Domba, Isolasi Sosial, Suara, Tingkah Laku, Kesejahteraan Hewan
ABSTRACT
Engeldal SEC, Subandriyo, Handiwirawan E, Noor RR. 2012. Effect of different levels of social isolation on the acoustical characteristics of sheep vocalization. JITV 18(3): 208-219. DOI: 10.14334/jitv.v18i3.323.
In farm animals, vocal analysis is accepted as a non-invasive method for assessing animal welfare in comparison to most physiological measurements. The aim of this study was to investigate the influence of different degrees of social isolation on the acoustic characteristics of stress-induced bleats from three different breeds of sheep. The experiment consisted of subjecting the animals to three different levels of social isolation during which both locomotive and vocal data were recorded. The data on locomotive behaviour was subjected to descriptive analysis while specialized acoustic software was used to measure a total of thirty-six parameters of recorded bleats. Descriptive analysis showed that the animals displayed more locomotive activity during partial isolation compared to complete isolation. Number of bleats during partial isolation was also found to be higher. The application of two-way analysis of variance showed a significant effect of isolation level and breed on both temporal and structural acoustic properties. Amplitude, power and time acoustic properties were found to affect acoustic quality of vocal responses to isolation, whereas frequency related properties were also found to be different significantly (P < 0.05) between breeds. From spectrogram analysis, the patterns of energy distribution within the calls proved to offer the most evident that differ between isolation levels and breeds. It was concluded that acoustic analysis can be helpful in revealing the affective state of socially isolated rams.
Key Words: Sheep, Social Isolation, Vocalization, Behaviour, Animal Welfare
INTRODUCTION
Acoustic communication has been found to play an important role in the social life of many animals. Communicative behaviour occurs in any form of social interaction and implies an exchange of information between at least two individuals: a signaler and a receiver. The acquisition and the use of information helps animals to anticipate and respond appropriately to events, and therefore to increase their survival (Owings et al. 1997).
According to De Lucia et al. (2010) vocalizations are essential in communication and social interactions, conveying the speaker's identity, gender, intentions, and emotional state. Dawkins (2004) suggested that vocalizations may perhaps be a rather special case of an indicator of what an animal wants because they are signals, that is, behaviours that have specially evolved to alter the behaviour of another animal, and which can therefore be "listened in on" by humans. In communication, information is made available by signals that vary in relation to the type of information delivered and to the surrounding environment (Vannoni et al. 2005).
The passing of more restrictive animal welfare laws has caused the detection of stress, especially in domesticated animals, to become an important issue. One suggested method of detecting stress has been through monitoring the vocalizations of the animals (Clemins et al. 2005). In sheep and goat, isolation stress resulted in increased vocalization and in variable and statistically nonsignificant increases in heart, pulse and respiratory rates. Isolation caused significant increases in the plasma concentrations of cortisol and glucose and a decrease in that of magnesium (Al-Qarawi and Ali 2005) and strees can affect livestock production. The vocalizations produced by an animal species can be categorized from both structural and functional standpoints, and the relationship between the two may take many forms as is believed by Soltis et al. (2005).
It is possible to assess welfare using some animal signals shown when they need certain resources. Weary and Fraser (1995) have reported that vocal and other natural signals provide reliable indicators of the signaller's needs. Acoustic signals can vary in their duration, pitch (vibration frequency), or amplitude. These features can be described and studied with the aid of spectrograms which give a visual representation of a sound (Hauser 1996).
Briefer (2012) believes that regardless of how the communication occurs, it is important to understand how animals can benefit from producing signals, and how others can benefit from responding. Bioacoustics is the study of sound in animals and includes, but is not limited to, animal communication with associated behaviour (Bradbury and Vehrencamp 1998), sound production anatomy and neurophysiology, auditory capacities and auditory mechanisms, and animal welfare (Manteuffel et al. 2004). The main goal of this field of study is to determine the role of animal vocalizations in the communication process. Knowledge of the value of vocal parameters may be of great importance within the field of animal production (Brudzynski 2010)
Animal acoustic behaviour can be termed as being rather complex due to its variability. Mammalian vocalization consists of a varied number of different call types and contexts in which these calls typically occur (Fitch et al. 2002). It is therefore of great importance to gain understanding in the process of vocal production. Taylor and Reby (2010) reported that physiological fluctuations in emotional or motivational state influence the acoustic characteristics of signals in a reliable and predictable manner. With only a mere few exceptions, mammal vocalizations are believed to be genetically determined with only limited flexibility (Seyfarth and Cheney, 2010). A careful description of the behaviour patterns or a sequence of behaviours of animals offers the possibility to identify all of the relevant components and to link their performance to the wider context of their physical and biological environment (Scott 2005).
Sheep are very gregarious animals and the isolation of an individual from its conspecifics leads to the manifestation of well defined distress behaviours (Poindron et al. 1997). According to Vannoni et al. (2005) distress calls are emitted by animals of this species during times of acute distress. The call is usually long, uncontrolled, very loud and given with the mouth open. According to the authors, distress calls are probably directed non-specifically and induce strong reactions in all conspecifics that become instantaneously and acutely alerted when they hear these sounds. Research of an acoustic features of stress-induced vocalizations cause of social isolation in a local Garut sheep and crossbred sheep was reported yet.
The purpose of this study was to analyze the acoustic features of stress-induced vocalizations of adult rams at different levels of social isolation to discover which acoustic measures may serve as an indicator of the animals' motivational state and distress level. The recorded data were also used to examine the difference between the characteristics of vocalizations based on breed. This was done to investigate the possibility that breeds of sheep may differ in their vocal response to stressful situations. Additionally, other behavioural data were recorded that might help define the animals' stress response.
MATERIAL AND METHODS
Study site and animals
The study was conducted at the Sheep Research Facility of the Indonesian Research Institute for Animal Production situated in Bogor. During the course of the study twelve randomly selected, clinically healthy, 2-3 year old adult rams were used. The animals were of three breeds, namely Barbados Blackbelly Cross (BC) (50% Local Sumatera, 50% Barbados Blackbelly), Local Garut (LG) and Composite Garut (KG) (50% Local Garut, 25% St. Croix, 25% Moulton Charollais).
Isolation procedure
Four animals from each breed were subjected to three individual tests for a maximum of 15 min. Each ram was subjected to isolation once per day on three consecutive days. Two persons drove each animal from the group pens to the testing room, which consisted of a 2.90 x 2.75 m enclosure with plain walls. The tests were as follows:
Test 1. Subject with human observer
This procedure consisted of catching the animal in its home pen and transporting it to the observation area where it was locked inside the enclosure together with the observer. During this procedure the animal had no visual or tactile contact with conspecifics. Olfactory and acoustic contact with conspecifics were, however, still possible.
Test 2. Subject completely alone
During this procedure the focal animal was transported to the observation area, where it was kept completely alone without visual or tactile contact with conspecifics. There was, however, still a possibility for olfactory and acoustic contact. During this procedure the animal's behaviour was observed through a small window while the observer remained hidden from the focal animal.
Test 3. Partial isolation
For this procedure group pens with a measurement of 5 x 2.6 m and slatted flooring were used. The focal animal was moved from its home pen and entered into an adjacent pen where it was held completely alone. This experiment allowed the isolated animal to maintain visual, acoustic and olfactory contact with conspecifics.
Table 1 gives an overview of the experimental design which was used in this study.
Recorded behaviour
During the isolation procedures the frequency of specific state – and point event behaviours was recorded following the method used by Siebert et al. (2011). The occurrence of each of these behaviours was recorded during 5 min per observation period for each focal animal. The ethogram consisted of the following behaviours:
1. Locomotion: the number of single steps made by the animal both forward and backward.
2. Standing: animal stands in one place for longer than 15 seconds.
3. Lying: the animal's abdomen contacts the floor of the experimental area.
4. Rearing: the animal raises both its forelegs to the wall or bars of the pen and stands on its hind legs.
5. Jumping: all of the animal's four legs are simultaneously raised of the floor.
6. Vocalizations: number of bleats uttered by the focal animal.
Vocal data collection and analysis
Sheep vocalizations were recorded using a handheld Digital Voice Recorder (CENIX Type W900). During recording, the voice recorder was mounted on a rod and maintained around two to three meters from the vocalizing animal. Sound files were saved in mp3 format after which they were transferred to a computer for subsequent treatment. From each ram three bleats with the highest quality were selected. To increase the quality of the vocal samples each was filtered using the software program Wavepad Sound Editor 5.10 to remove both noise and hiss sounds. To identify possible differences between the bleats uttered by the rams during the various isolation procedures, a total of 36 time, frequency, amplitude and power acoustic parameters were measured using the software program Raven Pro Ver. 1.4 created by the Cornell Lab of
Table 1. Experimental design of isolation procedures
Day |
||||
Test |
Breed |
1 |
2 |
3 |
Subject with human observer |
Barbados Blackbelly Cross |
4 |
||
Local Garut |
4 |
|||
Composite Garut |
4 |
|||
Subject completely alone |
Barbados Blackbelly Cross |
4 |
||
Local Garut |
4 |
|||
Composite Garut |
4 |
|||
Partial isolation |
Barbados Blackbelly Cross |
4 |
||
Local Garut |
4 |
|||
Composite Garut |
4 |
|||
Ornithology, USA. Measurements were taken from the entire element at the same scale on the screen and with standard spectral settings. Bleats were digitized and archived at a sampling rate of 32 kHz and 16-bit accuracy in WAV format. Table 2 shows a complete list of the 36 acoustic measures taken on each call as defined by Charif et al. (2008).
Table 2. Acoustic measures
Measures |
Definition |
Average Power (AvgPower) |
Value of the power spectrum averaged over the frequency extend of the selection (Units: dB) |
Energy |
Total amount of energy within the selection bounds (Units: dB) |
Aggregate Entropy (AggEntropy) |
Disorder in a sound by analyzing the energy distribution within a selection (Units: u) |
Max Frequency (MxFreq) |
Frequency at which Max Power occurs within the selection (Units: Hz) |
Peak Frequency (PFreq) |
Frequency at which Peak Power occurs within the selection (Units: Hz) |
Max Power/Peak Power (MxPower/PPower) |
Maximum power in the selection (Units: dB) |
Center Frequency (CenterFreq) |
Frequency that divides the selection into two frequency intervals of equal energy (Units: Hz) |
1st Quartile Frequency (Q1Freq) |
Frequency that divides the selection into two frequency intervals containing 25% and 75% of the energy in the selection (Units: Hz) |
3rd Quartile frequency (Q3Freq) |
Frequency that divides the selection into two frequency intervals containing 75% and 25% of the energy in the selection (Units: Hz) |
IQR (Inter-quartile range) Bandwidth (IQRBW) |
Difference between the 1st and 3rd Quartile Frequencies (Units: Hz) |
Frequency 5% (Freq5) |
Frequency that divides the selection into two frequency intervals containing 5% and 95% of the energy in the selection (Units Hz) |
Frequency 95% (Freq95) |
Frequency that divides the selection into two frequency intervals containing 95% and 5% of the energy in the selection (Units: Hz) |
Bandwidth 90% (BW90) |
Difference between the 5% and 95% frequencies (Units: Hz) |
Center Time (CenterTime) |
Point in time at which the selection is divided into two time intervals of equal energy (Units: seconds) |
1st Quartile Time (Q1Time) |
Point in time that divides the selection into two time intervals containing 25% and 75% of the energy in the selection (Units: seconds) |
3rd Quartile Time (Q3Time) |
Point in time that divides the selection into two time intervals containing 75% and 25% of the energy in the selection (Units: seconds) |
IQR (Inter-quartile Range) Duration (IQRDur) |
Difference between the 1st and 3rd Quartile Times (Units: seconds) |
Time 5% (Time5) |
Point in time that divides the selection into two time intervals containing 5% and 95% of the energy in the selection (Units: seconds) |
Time 95% (Time95) |
Point in time that divides the selection into two time intervals containing 95% and 5% of the energy in the selection (Units: seconds) |
Duration 90% (Dur90) |
Difference between the 5% and 95% times (Units: seconds) |
Max Amplitude (MxAmpl) |
Maximum of all the sample values in the selection (Units: dimensionless sample values) |
Min Amplitude (MinAmpl) |
Minimum of all sample values in the selection (Units: dimensionless sample values) |
Peak Amplitude (PAmpl) |
Greater of the absolute values of max Amplitude and Min Amplitude (Units: dimensionless) |
RMS Amplitude (RMSAmpl) |
Root-mean-square amplitude of the selected part of the signal (Units: dimensionless sample units) |
Filtered RMS Amplitude (FRSAmpl) |
Root-mean-square amplitude for the selected time and frequency range (Units: dimensionless sample units) |
Max Time (MxTime) |
First time in the selection at which a sample with amplitude equal to Max Amplitude occurs (waveform view). For a spectrogram view, the first time in the selection at which a spectrogram point with power equal to Max Power/Peak Power occurs (Units: seconds) |
Min Time (MinTime) |
First time in the selection at which a sample with amplitude equal to Min amplitude occurs (units: seconds) |
Continued from Table 2 |
|
Measures |
Definition |
Peak Time (PTime) |
First time in the selection at which a sample with amplitude equal to Peak Amplitude occurs (Units: seconds) |
Begin Time (BTime) |
Time at which the selection begins (Units: seconds) |
Delta Frequency (DeltaFreq) |
Difference between the upper and the lower frequency limits of the selection (Units: Hz) |
Delta Time (DeltaTime) |
Difference between Begin Time and End Time for the selection (Units: seconds) |
End Time (ETime) |
Time at which the selection ends (Units: seconds) |
High Frequency (HighFreq) |
Upper frequency bound of the selection (Units: Hz) |
Low Frequency (LowFreq) |
Lower frequency bound of the selection (Units: Hz) |
File Offset (FOffset) |
Amount of time between the beginning of the file and the start of the selection (Units: seconds) |
Length |
Number of frames contained in a selection (Units: frames) |
Statistical analysis
The overall results for breeds and isolation level over the three days were descriptively analyzed using the program package SPSS 16.0 for Windows (SPSS Inc., Chicago, IL, USA). An average value for each acoustic trait for each individual was included in the statistical analysis. The data were tested for homoscedasticity and normality before using parametric tests. Two-way analysis of variance (ANOVA) was used for the assessment of effects due to isolation level and breed on ram vocalization. In all cases, a P < 0.05 was considered statistically significant. Duncan test was applied for post hoc comparison. Statistical tests were performed using the statistical package SAS for Windows, release 9.2 (SAS Institute, Inc., 2009). A General Linear Mixed model (GLM) analysis was used to investigate the effect of breed and level of isolation on the measured acoustic parameters. This was done using the following basic model:
Yijk = μ + Bi + Ij + (BI)ij + εijk
1. Yijk is the observation of the i-th breed of the j-th degree of social isolation, and the k-the replication
2. μ is the population mean
3. Bi is the effect of the i-th breed (i = 1, 2, 3)
4. Ij is an effect of the j-th degree of social isolation (j = 1, 2, 3)
5. BIij is an interaction between the effect of the i-th breed and the effect of the j-th degree of isolation
6. εijk is a random error due to the effect of the i-th breed, the j-th degree of social isolation, and the k-th is a replication
RESULTS AND DISCUSSION
Behavioural parameters
Sharp et al. (2002) reported that social isolation represents a stressful challenge to gregarious species, which is thought to negatively affect their welfare. Enhanced general activity and an increased level of vocalization are the initial responses that have been described for a number of farm animal species in social isolation, including cattle (Boissy and Le Neindre 1997), sheep and goats (Baldock and Sibly 1990), pigs (Schrader and Ladewig 1999; Weary and Fraser 1995) and chicks (Marx et al. 2001). The frequencies of the behaviours observed at the different levels of social isolation are represented in Table 3.
Table 3 Frequencies of observed behaviours at different levels of isolation
Isolation level |
||
Behaviour |
Complete |
Partial |
Locomotor activity |
90.63 ± 36.27 |
104.00 ± 49.25 |
Standing |
18.88 ± 5.77 |
13.92 ± 5.14 |
Lying |
0.00 ± 0.00 |
0.00 ± 0.00 |
Rearing |
1.04 ± 2.69 |
6.42 ± 8.09 |
Jumping |
0.21 ± 0.58 |
0.00 ± 0.00 |
Vocalization |
18.75 ± 11.89 |
46.17 ± 9.82 |
Values are means ± S.D.
Locomotor activity and vocalization were observed at higher frequencies in reacting animals during partial isolation compared to complete isolation. These results
are in line with those found by Siebert et al. (2011) for dwarf goats. In their study it was found that different degrees of isolation were clearly related to different behavioural and vocal responses, with higher rates of locomotion and high bleats compared to the completely isolated animals. Their results also showed higher rates of rearing and jumping for partially isolated animals compared to the animals which were completely isolated. The results of this study only partly support those findings. The mean values for lying were all found to be equal to zero, meaning that the animals did not show this behaviour at all during the observation periods. Partially isolated rams were, however, found to display rearing behaviour at a higher frequency as compared to when the animals were completely isolated. Baldock and Sibly (1990) and Carbonaro et al. (1992) also reported that social isolation induces a strong behavioural response in the form of enhanced locomotion and increased vocalization in both sheep and goats.
According to Herskin and Jensen (2000) the impact of social isolation on behaviour is affected by the level of contact (olfactory, visual, acoustic, social) that the isolated animals are allowed to maintain with their conspecifics. Based on their findings Siebert et al. (2011) have suggested that in partial isolation the permanent sensory feedback from conspecifics located in adjacent pens, may facilitate an active response pattern, increase behavioural arousal and support socially motivated efforts to restore social contact in partially isolated animals. Degree of isolation affected whether the animals displayed more active or inactive behaviours. Rams were observed standing more frequently when completely isolated compared to when partially isolated. This response could possibly be explained by the fact that the animals were more motivated to attempt to reunite with conspecifics when these were in sight, thus causing them to move around more in an attempt to escape.
Vocalization and call measures
Comparative studies on the vocalization of humans and animals have shown that structural and temporal variations in communication sounds serve several functions, such as to reliably transmit the affective state and individuality of the sender (Schehka et al. 2007). Results of the vocal analysis are shown in Table 4 and Table 5. Representative records of the acoustic parameters of bleats at different levels of social isolation are given in Table 4 while the results from the vocal analysis of bleats from different breeds of sheep are shown in Table 5. Included in these two tables are only those measurements which showed significant differences between the different groups.
Table 4. Comparison of acoustic parameters of high-pitched bleats at different levels of isolation
Level of isolation |
|||
Acoustic parameters |
Partial |
Completely alone |
With observer |
Amplitude |
|||
MxAmpl (u) |
13690.0b ± 2848.2 |
27381.0a ± 4999.9 |
26784.0a ± 5128.1 |
MinAmpl (u) |
-12874.0a ± 3574.8 |
-27560.0b ± 5200.6 |
-26777.0b ± 4814.1 |
PAmpl (u) |
14141.0b ± 3173.4 |
27902.0a ± 4880.2 |
27576.0a ± 4613.2 |
RMSAmpl (u) |
2551.3b ± 941.3 |
6399.5a ± 1784.7 |
5867.1a ± 1687.4 |
FRSAmpl (u) |
2551.3b ± 941.3 |
6399.4a ± 1784.7 |
5867.1a ± 1687.4 |
Power |
|||
MxPower (dB) |
96.0b ± 3.3 |
103.6a ± 2.3 |
103.1a ± 2.3 |
PPow (dB) |
96.0b ± 3.3 |
103.6a ± 2.3 |
103.1a ± 2.3 |
AvgPower (dB) |
70.3b ± 3.5 |
78.7a ± 2.5 |
77.6a ± 2.5 |
Energy (dB) |
113.4b ± 3.5 |
121.7a ± 2.9 |
121.0a ± 2.9 |
Time |
|||
PTime (s) |
0.6b ± 0.2 |
0.7ab ± 0.2 |
0.8a ± 0.3 |
Time5 (s) |
0.4a ± 0.1 |
0.3b ± 0.1 |
0.4ab ± 0.1 |
Values are least square means ± S.D.
a,b Different letters in the same row indicate significant difference between groups (P < 0.05)
MxAmpl |
= |
Max Amplitude, |
MinAmpl |
= |
Min Amplitude |
PAmpl |
= |
Peak Amplitude, |
RMSAmpl |
= |
Root-square-mean Amplitude |
PPower |
= |
Peak Power, |
AvgPower |
= |
Average Power |
PTime |
= |
Peak Time, |
Time5 |
= |
Time 5% |
FRSAmpl |
= |
Filtered root-square-mean Amplitude, |
MxPower |
= |
Max Power |
There was substantial variation in the acoustic measures of high pitched bleats from partially and completely isolated rams. Brudzynski (2010) believes that most mammalian species use both changes of acoustic sound parameters as well as temporal patterning in coding their signals. The results from the vocal analysis showed significant differences in both temporal (Time) and structural (Amplitude, Power) parameters for calls uttered during different levels of social isolation (Table 4). Significant differences were found for the values of Max Amplitude (MxAmpl), Peak Amplitude (PAmpl) Root-mean-square Amplitude (RMSAmpl) and Filtered root-mean-square Amplitude (FRSAmpl), with all of these parameters being higher in bleats uttered during complete isolation as compared to bleats from partially isolated animals. Calls with the highest Max Amplitude (MxAmpl) were found for animals which were completely isolated. These results may thus indicate that the intensity of vocalizations (high amplitude) uttered by rams increases with the stressfulness of a specific situation.
Bleats from completely isolated animals all had a higher Power and Energy level which was found to be significantly different (P < 0.05) from the Power and Energy content from bleats uttered during partial isolation. Calls with the highest Peak Time (PTime), were found during complete isolation. Time 5% (Time5) was found to be highest for calls from partially isolated animals. These results thus reveal that more effort was put into calls made during complete isolation as compared to those made during partial isolation. There were no significant differences found for any of the measured parameters for calls from rams which were kept completely alone and the ones which were in the presence of the observer. It showed that presence of observer cannot make the sheep calm or two treatment conditions (completely alone and with observer) are the same for the sheep. Siebert et al. (2011) found that high bleats from goats that were completely isolated had lower 3rd Quartile and Entropy compared to those in partial isolation. In this study, however, the impact of isolation on 3rd Quartile and Entropy did not indicate a clear tendency in any direction in either degree of isolation.
All of the calls used for analysis were high-pitched bleats, meaning that they were characterized by high frequencies and made with the mouth open as defined by Siebert et al. (2011). Weary and Fraser (1995), based on their study with piglets, reported that high-pitched isolation calls can be considered part of an honest signaling system that reflects the level of need and the state of welfare of the sender. In this study the isolated rams only emitted high-pitched bleats during the observation sessions. This finding corresponds well with the findings of Siebertet al. (2011) who found that socially isolated goats emitted twice the amount of high bleats compared to the amount of low bleats. They mentioned that high pitched bleats could be assumed to be honest but costly signals that communicate the need of isolated animals to restore social relationships. This relates well to the results found during this study, where the high-pitched bleats uttered during complete isolation were found to contain the highest amount of energy. Due to the higher level of energy found in high-pitched bleats, these types of vocalizations are generally considered to be costly signals that are related to a high level of excitement and distress (Puppe et al. 2005).
Breed differences of acoustic structure of high bleats
Twenty-three of the 36 parameters that were measured from the bleat recordings differed significantly between the three sheep breeds. Of the temporal parameters, Max Time (MxTime), Min Time (MinTime), Center Time (CenterTime), Peak Time (PTime), Time 5% (Time5), Time 95% (Time95), 1st Quartile Time (Q1Time) and 3rd Quartile Time (Q3Time) were all found to be lowest for the breed Local Garut (Table 5) and to differ significantly from both Barbados Blackbelly Cross and Composite Garut. The duration of calls was not found to differ significantly between breeds nor isolation level.
Of the parameters included in the Amplitude group MxAmpl, PAmpl, RMSAmpl, FRSAmpl were found to be highest for the breed Barbados Blackbelly Cross. The amplitude of an acoustic signal is believed to be a key factor for the exchange of information, determining the broadcast area, or active space, of the signal according to Gil and Gahr (2002). Average Entropy (AvgEntropy) was highest in bleats from Composite Garut and lowest for the breed Barbados Blackbelly Cross.
All frequency-related parameters (Q1Freq, Q3Freq, BW90, Freq95, IQRBW) were found to be highest for the breed Composite Garut and lowest for the breed Barbados Blackbelly Cross. The values of these parameters were not found to differ significantly for the breed Local Garut compared to those from both Barbados Blackbelly Cross and Composte Garut. The Peak Frequency of the high-pitched bleats did not differ significantly between breeds (P ˃ 0.05), but animals from the breed Composite Garut responded to isolation with bleats with the highest overall frequencies (Table 5).
Max Power, Peak Power, Average Power and Energy, were all found to be highest in bleats from Barbados Blackbelly Cross rams. These parameters did not differ significantly in the calls from Local Garut and Composite Garut.
Sound production by animals is often based on complex mechanical events, ranging from cavitation induced sounds by shrimps to the energy conversion of flow to vibrating structures in the larynx of mammals (Versluis et al. 2000).The visual inspection of the spectrograms of which examples are given in Figures 1, 2 and 3, underlines a difference in the energy distribution in bleats uttered during both partial and complete isolation. The spectrograms from bleats made during complete isolation show a higher amount of energy as compared to bleats made during partial isolation. Application of two-way ANOVA confirmed the results obtained from visual inspection of spectrograms. Significant differences were found among the three breeds at the different levels of social isolation in the amount of energy included in the calls (Table 5).
Table 5. Comparison of acoustic parameters of high-pitched bleats from different breeds
Breed |
|||
Acoustic parameters |
BC |
LG |
KG |
Amplitude |
|||
MxAmpl (u) |
26263.0a ± 7891.0 |
21239.0b ± 6174.5 |
20354.0b ± 8215.7 |
PAmpl (u) |
26785.0a ± 7752.3 |
21906.0b ± 6359.5 |
20927.0b ± 8226.7 |
RMSAmpl (u) |
6368.2a ± 2379.6 |
4184.7b ± 1557.4 |
4265.0b ± 2229.8 |
FRSAmpl (u) |
6368.2a ± 2379.6 |
4184.6b ± 1557.3 |
4265.0b ± 2229.8 |
AvgEntropy (u) |
2.9b ± 0.2 |
3.1ab ± 0.2 |
3.2a ± 0.2 |
Power |
|||
MxPower (dB) |
103.0a ± 4.0 |
100.2b ± 3.7 |
99.4b ± 4.7 |
PPow (dB) |
103.0a ± 4.0 |
100.2b ± 3.7 |
99.4b ± 4.7 |
AvgPower (dB) |
78.3a ± 3.9 |
74.6b ± 3.8 |
73.7b ± 5.2 |
Energy (dB) |
121.8a ± 4.2 |
117.3b ± 3.5 |
117.0b ± 5.5 |
Time |
|||
MxTime (s) |
0.8a ± 0.2 |
0.5b ± 0.2 |
0.7a ± 0.2 |
MinTime (s) |
0.7a ± 0.2 |
0.5b ± 0.1 |
0.7a ± 0.2 |
CenterTime (s) |
0.8a ± 0.2 |
0.6b ± 0.1 |
0.8a ± 0.2 |
PTime (s) |
0.7a ± 0.1 |
0.5b ± 0.1 |
0.7a ± 0.2 |
Time5 (s) |
0.4a ± 0.1 |
0.3b ± 0.1 |
0.4a ± 0.1 |
Time95(s) |
1.2ab ± 0.3 |
1.0b ± 0.2 |
1.3a ± 0.3 |
Q1Time (s) |
0.6a ± 0.1 |
0.5b ± 0.1 |
0.6a ± 0.1 |
Q3Time (s) |
1.0a ± 0.2 |
0.8b ± 0.1 |
1.0a ± 0.2 |
Frequency |
|||
Q1Freq (Hz) |
670.1b ± 69.9 |
718.8ab ± 71.3 |
732.6a ± 65.2 |
Q3Freq (Hz) |
1236.1b ± 307.9 |
1375.0ab ± 288.1 |
1531.2a ± 170.5 |
BW90 (Hz) |
1420.1b ± 292.7 |
1576.4ab ± 208.6 |
1756.9a ± 310.2 |
Freq95 (Hz) |
1871.5b ± 286.7 |
2066.0ab ± 177.2 |
2257.0a ± 219.6 |
IQRBW (Hz) |
566.0b ± 269.1 |
656.2ab ± 243.7 |
798.6a ± 170.3 |
Values are least square means ± S.D.
a,b Different letters in the same row indicate significant difference between groups (P < 0.05)
Breed: BC |
= |
Barbados Blackbelly Cross, |
LG |
= |
Local Garut, |
KG |
= |
Composite Garut |
MxAmpl |
= |
Max Amplitude, |
MinAmpl |
= |
Min Amplitude, |
PAmpl |
= |
Peak Amplitude. |
RMSAmpl |
= |
Root-square-mean Amplitude, |
FRSAmpl |
= |
Filtered root-square-mean Amplitude, |
AvgEntropy |
= |
Average Entropy, = |
MxPower |
= |
Max Power, |
PPower |
= |
Peak Power, |
AvgPower |
= |
Average Power, |
MxTime |
= |
Max Time, |
MinTime |
= |
Min Time, |
CenterTime |
= |
Center Time, |
PTime |
= |
Peak Time, |
Time5 |
= |
Time 5%, |
Time95 |
= |
Time 95%, |
Q1Time |
= |
1st Quartile Time, |
Q3Time |
= |
3rd Quartile Time, |
Q1Freq |
= |
1st Quartile Frequency, = |
Q3Freq |
= |
3rd Quartile Frequency, |
BW90 |
= |
Bandwidth 90%, |
Freq95= |
= |
Frequency 95%, |
IQRBW |
= |
Inter-quartile-range bandwidth |
Kendrick (2008) reported that there are significant differences in the fundamental spectrographic patterns between individual animals. Thus animals should be able to be identified based on their high-pitch bleats. According to the author the sound spectrogram of the same sheep producing a high pitch bleat, when it is excited as opposed to stressed or fearful, is also clearly different. When the vocalization is produced in a positive state of excitement it uses a broader frequency range, especially at the higher frequencies and shows distinct regular bands of alternating high and low intensity. When it is produced during stressful circumstances it is thought to exhibit less representation in the higher frequency ranges and almost completely loses the bands of intensity modulation. The results from this study, however, did not show any significant differences in frequency range of bleats at different levels of social isolation nor for the different breeds. Based on the above mentioned statements, however, it can be understood that one call, based on its acoustic content, may indeed be able to communicate different things to other sheep in different circumstances.
The results from Siebertet al. (2011) showed that compared to partially isolated individuals, goats in complete isolation emitted high bleats with more pronounced pureness (lower values of 3rd Quartile) and diminished randomness (lower values of Entropy). According to them, reduced sound variability might be viewed as a consequence of higher distress that could be related to changes in psycho-physiological mechanisms of sound production in the vocal tract.
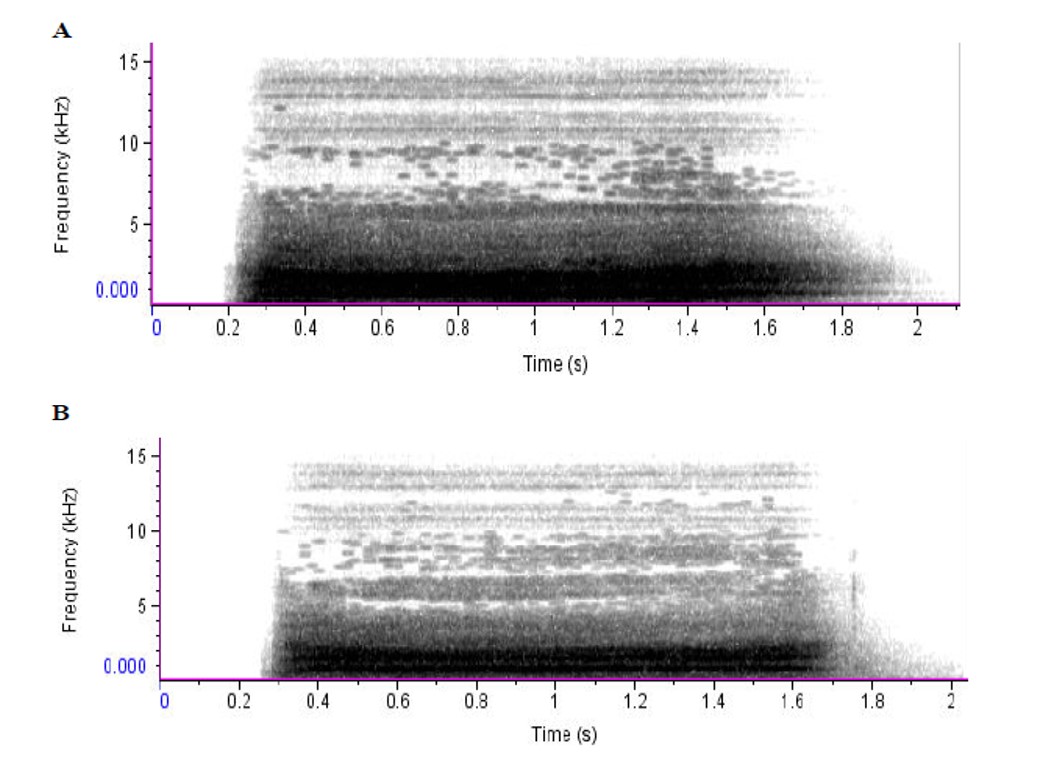
Figure 1. Representative spectrographs of high pitched bleats from the same Barbados Blackbelly Cross ram while (A) completely alone and (B) partially isolated. The dark grey areas represent large concentrations of sound energy in specific time and frequency. Parameters of the spectrograph included: FFT length of 256, and a bandwidth of 180 Hz. Hann window with overlap of 50%
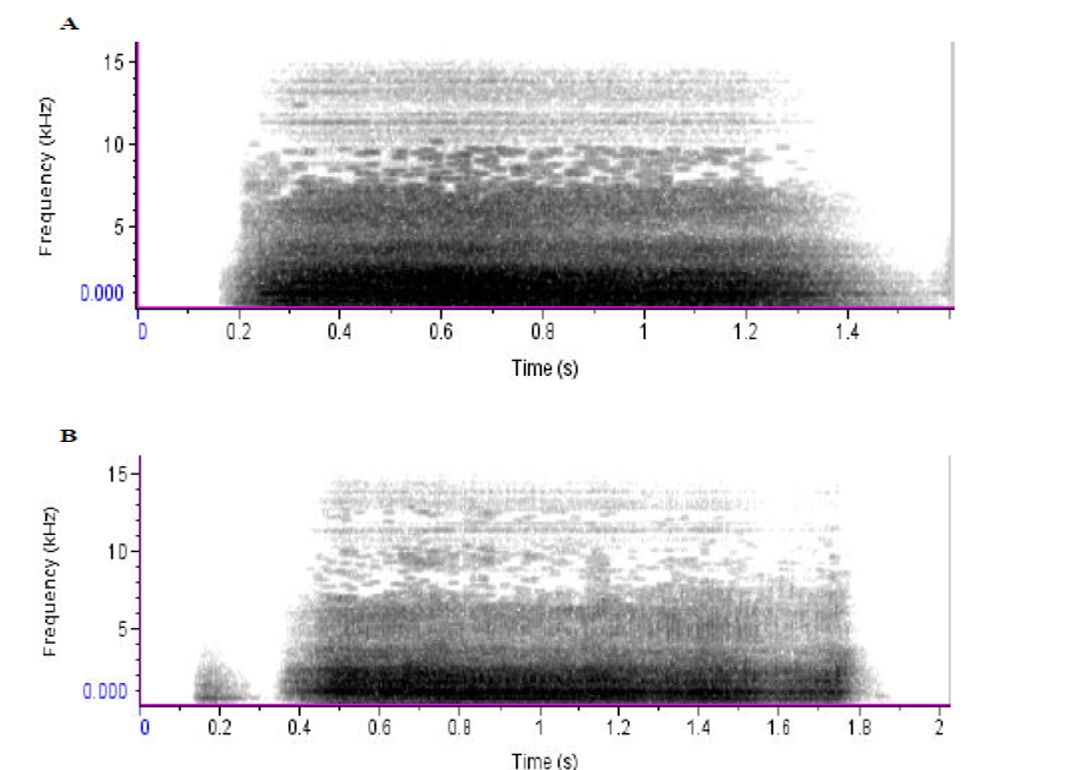
Figure 2. Representative spectrographs of high pitched bleats from the same Local Garut ram while (A) completely alone and (B) partially isolated. The dark grey areas represent large concentrations of sound energy in specific time and frequency. Parameters of the spectrograph included: FFT length of 256, and a bandwidth of 180 Hz. Hann window with overlap of 50%.
Figure 3. Representative spectrographs of high pitched bleats from the same Composite Garut ram while (A) completely alone and (B) partially isolated. The dark grey areas represent large concentrations of sound energy in specific time and frequency. Parameters of the spectrograph included: FFT length of 256, and a bandwidth of 180 Hz. Hann window with overlap of 50%
Similar results were found by Puppe et al. (2005), who reported that piglets responded with similar changes in these call measures during the surgical period of the painful castration process. In this study, it was found that the calls from Composite Garut rams had the highest level of Entropy indicating that bleats uttered by this breed exhibit an increased level of randomness compared to that of the other two breeds. The results also show the highest Q3 value for bleats from
Garut, which indicates that their vocalizations contain less pronounced pureness. Based on the statements by Siebert et al. (2011), these findings thus suggest that the bleats from these animals reveal a lower level of distress.
CONCLUSIONS
Based on the results obtained in this study it can be concluded that different levels of social isolation induce different behavioural and vocalization responses in rams from different breeds. Amount of noise energy distributed throughout the call was proven to be the most discriminant vocal characteristic based on degree of social isolation and breed. It is believed that these results provide support that specific measures of vocal quality may indeed serve as a valuable tool for evaluating affective-related behaviour and thus also welfare status in adult rams.
For stress reducing and importance of animal welfare, sheep should be housed together with the other sheep or one of alternative if the condition not possible can be housed partially isolation. Stress due to isolation in the long term can affect the production of sheep. Increased production of sheep is a reflection of the attention to animal welfare and reduces of livestock stress.
REFERENCES
Al-Qarawi AA, Ali BH. 2005. Isolation stress in desert sheep and goats and the influence of pretreatment withxylazine or sodium betaine. Vet Res Commun. 29:81-90.
Baldock NM, Sibly RM. 1990. Effects of handling and transportation on the heart rate and behaviour of sheep. Appl Anim Behav Sci. 28:15-39.
Boissy A, Le Neindre P. 1997. Behavioral, cardiac and cortisol responses to brief peer separation and reunion in cattle. Phys Behav. 61:693-699.
Bradbury JV, Vehrencamp SL. 1998. Principles of animal communication. Sunderland: Sinauer Associates, Inc.
Briefer EF. 2012. Vocal expression of emotions in mammals: mechanisms of production and evidence. J Zool. 14:79-98.
Brudzynski SM. 2010. Handbook of mammalian vocalization: An integrative neuroscience approach. Oxford (UK): Academic Press.
Carbonaro DA, Friend TH, Dellmeier GR, Nuti LC. 1992. Behavioural and physiological responses of dairy goats to isolation. Physiol Behav. 51:297-301.
Charif RA, Waack AM, Strickman LM. 2008. Raven Pro 1.3 User's Manual. Ithaca (NY): Cornell Laboratory of Ornithology.
Clemins PJ, Johnson MT, Leong KM, Savage A. 2005. Automatic classification and speaker identification of African elephant (Loxodonta africana). J Acous Soc Amer. 177:956-963.
Dawkins MS. 2004. Using behaviour to assess animal welfare. Anim Welfare. 13:3-7.
De Lucia M, Clarke S, Murray MM. 2010. A temporal hierarchy for conspecific vocalization discrimination in humans. J Neuro Sci. 33:11210-11221.
Fitch TW, Neubauer J, Herzel HP. 2002. Calls out of chaos: the adaptive significance of nonlinear phenomena in mammalian vocal production. Anim Behav. 63:407-418.
Gil D, Gahr M. 2002. The honesty of bird song: multiple constraints for multiple traits. Trends Ecol Evol. 17:133-141.
Hauser MD. 1996. The evolution of communication. Massachusetts: MIT Press.
Herskin MS, Jensen KH. 2000. Effects of different degrees of social isolation on the behaviour of weaned piglets kept for experimental purposes. Anim Welfare.9:237-249.
Kendrick KM. 2008. Sheep senses, social cognition and capacity for consciousness. In: Dwyer CM, editor. The welfare of sheep. Cambridge (UK): Springer Science + Business Media B.V. p. 135-139.
Manteuffel G, Puppe B, Schon PC. 2004. Vocalization of farm animals as a measure of welfare. Appl Anim Behav Sci. 88:163-182.
Marx G, Leppelt J, Ellendorff F. 2001. Vocalisation in chicks (Gallus gallus dom.) during stepwise social isolation. Appl Anim Behav Sci. 75:61-74.
Owings DH, Beecher M, Thompson N. 1997. Communication perspectives in ethology. Vol12. New York: Plenum Press.
Poindron P, Soto R, Romeyer A. 1997. Decrease of response to social separation in preparturient ewes. Behav Process. 40:45-51.
Puppe B, Schön PC, Tuchscherer A, Manteuffel G. 2005. Castration-induced vocalisation in domestic piglets, Sus crofa: complex and specific alterations of the vocal quality. Appl Anim Behav Sci. 95:67-78.
Schehka S, Esser KH, Zimmermann E. 2007. Acoustical expression of arousal in conflict situations in tree shrews (Tupaia belangeri). J Comp Physiol A. 193:845-852.
Schrader L, Ladewig J. 1999. Temporal differences in the responses of the pituitary adrenocortical axis, the sympathoadrenomedullar axis, heart rate, and behaviour to a daily repeated stressor in domestic pigs. Physiol Behav. 66:775-783.
Scott G. 2005. Essential animal behavior. Victoria (Aust): Blackwell Publishing Ltd.
Seyfarth RM, Cheney DL. 2010. Production, usage, and comprehension in animal vocalizations. Brain Lang. 115:92-100.
Sharp JL, Zammit TG, Azar TA, Lawson DM. 2002. Stress-like responses to common procedures in male rats housed alone or with other rats. J Am Assoc Lab Anim Sci. 41:8-14.
Siebert K, Langbein J, Schön P, Tuchscherer A, Puppe B. 2011. Degree of social isolation affects behavioural and vocal response patterns in dwarf goats (Capra hirscus). J Appl Anim Behav Sci. 131:53-62.
Soltis J, Leong K, Savage A. 2005. African elephant vocal communication I: antiphonal calling behaviour among affiliated females. Anim Behav. 70:579-589.
Taylor AM, Reby D. 2010. The contribution of the source-filter theory to mammal vocal communication research. J Zool. 280:221-236.
Vannoni E, Torriani MVG, McElligott AG. 2005. Acoustic signaling in cervids: A methodological approach for measuring vocal communication in fallow deer. Zurich (Switzerland): Zoologisches Institut, Universitat Zurich.
Weary DM. Fraser D. 1995. Signalling need. Costly signals and animal welfare assessment. Appl Anim Behav Sci. 44:159-169.
TweetRefbacks
- There are currently no refbacks.

This work is licensed under a Creative Commons Attribution 4.0 International License.

























